- Mutazioni genetiche: scoperte per la prima volta da Hugo de Vries, consistono nella modificazione del genoma (maggiori informazioni).
- Dominanza incompleta: fenomeno che avviene quando, esaminando un carattere di una specie, gli individui eterozigoti non presentano il fenotipo dominante, come affermava Mendel, e tantomeno quello recessivo, bensì un nuovo carattere fenotipico "intermedio".

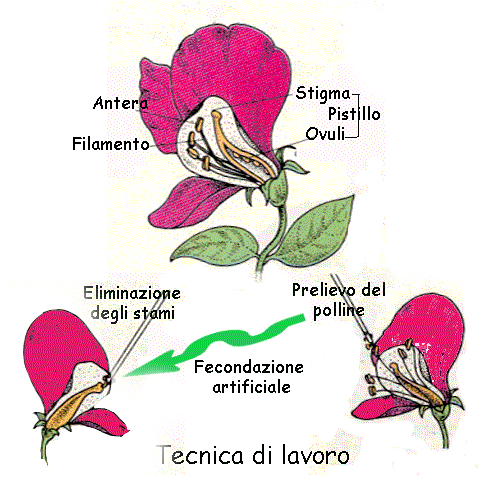
Il caso della "bocca di leone"
- Alleli multipli: se un carattere di una popolazione di individui è determinato da un gene che presenta più di due forme alleliche differenti.
- Codominanza: quando negli individui eterozigoti per un certo carattere i due alleli si manifestano contemporaneamente nei loro fenotipi omozigoti.

Il caso dei gruppi sanguigni umani, gene dagli alleli multipli: A e B sono dominanti allo stesso modo e 0 è recessivo; gli eterozigoti A0 e B0 manifesteranno rispettivamente il fenotipo A e B mentre gli eterozigoti AB manifestano sia A che B allo stesso modo, ossia il gruppo sanguigno AB.
- Epistasi: è quel fenomeno dove il particolare genotipo di un carattere dell'organismo inibisce l'espressione di altri geni.
- Eredità poligenica: i casi in cui un determinato carattere fenotipico di una specie è il risultato delle azioni combinate di due o più geni. Questi caratteri, detti appunto poligenici, si manifestano in genere con lievi differenze fenotipiche tra gli individui: si parla di variazione continua.
La tipica curva della variazione continua: nonostante possano variare i valori, l'andamento rimane sempre "a campana" perchè la maggior parte dei fenotipi sono molto simili.
- Pleiotropia: quei casi dove, invece, il genotipo di un singolo gene influisce su più caratteri fenotipici di un organismo.
- Ambiente: ogni carattere fenotipico di un individuo è il risultato dell'interazione tra il genotipo dello stesso e le condizioni ambientali a cui questo è sottoposto (nel senso che il suo fenotipo può variare a seconda dell'ambiente in cui esso si trova).

I colori dei fiori della primula variano a seconda della temperatura ambiente: le primule a fiori rosa o rossi sono tipiche nelle montagne ad alta quota mentre i fiori bianchi si trovano in ambienti più caldi.